Ward Laboratories: Regenerative Agriculture in context
Willie Pretorius, Soil Health Consultant at Ward Laboratories, explains Regenerative Agriculture in context, viz-a-viz the provision and utilization of natures provided resources.

Asking the age-old question of how does nature provide for the natural forests, grasslands and Prairies in such abundance and why is this not incorporated into our agricultural production systems? Why is this not happening at a large scale? We know that this is the basis for regenerative agriculture already embraced by many farmers around the world that have started a new industry within modern agriculture gaining traction and growing at a substantial rate.
Let us address the first issue of the abundant available resources that nature employs to maintain the natural vegetation and all other living entities and then ask the question why are the same resources not applied in commercial crop and animal production?
Let us dissect the building blocks of living entities, how these building blocks are assembled and the components of these building blocks.
All living entities from the most minute microbe to humans contain the following basic elements, C, H, O, N, P, K, Ca, Mg, S, Cl, Na, Fe, Mn, Cu, Zn, B, Mo, Ni. These are the essential nutrients for plant growth. However, many Scientists also include these elements in the plant; Se, Cr, Co, Ti, Sr, Sn, As, Ba, Ag, Zr. This would bring the number of elements to 28 grouped into macro, micro and nano required minerals. There are very respected scientists such as Dr. David Johnson who puts the number of essential elements at 87. For the purposes of this discussion we will keep to the number of 28 and point out what the consequences are for plant, animal and human health should these nutrients be required using the conventional adopted crop production system.
Where do these elemental building blocks originate from in nature’s supply model for the forests, grasslands, prairies, and other natural growing areas?
Elements originating from the atmosphere.
Oxygen (65% O), Hydrogen (10% H), Carbon (18% C), and Nitrogen (3% N) make up about 96% of the total minerals contained in living cells. Hydrogen and Oxygen are regarded as emanating from the atmosphere as water vapor and rain as H2O. Nitrogen in nature’s model emanates from the atmosphere and will be elaborated in more detail. The fact that the atmosphere consists of 78% Nitrogen gas fits with nature’s supply model.
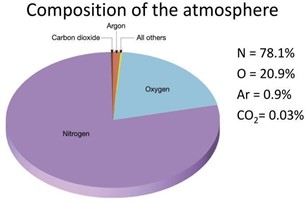
CO2 is now about 0.04%
Minerals originating from the soil make up about 4% of the total living cell mass.
P, K, Ca, Mg, S, Cl, Na, Fe, Mn, Cu, Zn, B, Mo, Ni, Se, Cr, Co, Ti, Sr, Sn, As, Ba, Ag, Zr
Macro, Micro, Nano
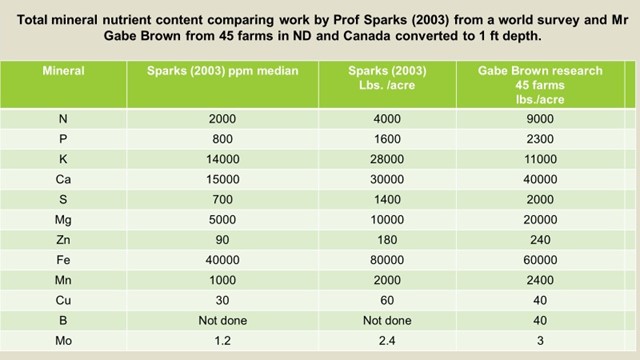
If we analyze soils on a complete basis using the “Total Nutrient Digestion” or “Total Nutrient
Extraction” (TNE) that analyzes not only the nutrients in suspension or those adhered to the cation exchange points as done in a soil chemical analyses, we get a very different picture of the actual mineral contents not necessarily available but potentially available.
Work done in this instance by Dr. Sparks from University of Delaware on averaging world soils is supported by work in progress by Mr. Gabe Brown for General Mills in ND and Canada.
Nature has to its disposal quanta of mineral nutrients albeit in an unavailable solid-state form as well as atmospheric minerals albeit in the gas form.
How does nature put its growth model together? With life, microbial life!!
In this instance it is important to understand that all living species have an inter dependence with microbial life, so much so that it is calculated that a human being contains about 100 trillion cells of which 90 million are microbial living in mutual association with the human cells. This mutual inter dependence forms the basis for the maintenance of plant, animal, and human health.
Describing nature’s model.
Carbon, C; Hydrogen, H; and Oxygen, O.
The process of photosynthesis through the absorption of CO2 using sunlight energy capture takes care of the carbon and partly of the oxygen building process into carbohydrates. The water absorption through roots takes care of the hydrogen and the other part of the oxygen incorporation into plants. During photosynthesis two thirds of the Oxygen returns to the atmosphere.
Nitrogen, N.
Nitrogen is probably the most difficult element to get an understanding of because it is potentially provided from two sources: the atmosphere and the soil. The atmosphere contains 78% N2 (Nitrogen gas) which is an inert gas with a very strong triple molecular bond, binding the gas in the double N gaseous state.
Soil also contains around 2000 to 5000 ppm of organic Nitrogen as can be seen from the table above.
From the Atmosphere.
Atmospheric nitrogen gas is fixed through a reductive biochemical process into an organic molecule as Ammonia in the presence of the enzyme nitrogenase facilitated by a specialized group of bacteria known as Diazotrophs in the first step of this process. This process is the same where the facilitation is through legume associated Rhizobium bacteria or through free living nitrogen fixing bacteria that live in association with grasses or other plant species. The next step is the binding of the Ammonia with organic acids to form amino acids which are the building blocks of proteins. In this instance it is important to understand that the amino acids are provided into the nutrient pool around the rhizosphere as plant or bacterial food as we call it in Soil Health terms: “water extractable organic nitrogen” (WEON). Plants absorb these amino acids as the precursor building blocks of proteins as do microbes.
There is strong evidence that supports the demand for nitrogen as for all other required nutrients within a plant is regulated through signaling systems between plant, bacteria, and fungi in this closely connected and linked system. That signals from the plant requiring a certain quantity of nitrogen be sent through to the roots where the nitrogen fixing bacteria are located within the rhizosphere as free livings or root associated with nodules in legumes, together with the sugars that feed this group of microbes so their numbers will multiply into a quorum for providing the nitrogen fixing service through the atmospheric nitrogen draw down into amino acids as already described. Once the tasks have been completed the food source is withdrawn and the excessive microbial population becomes a food source down the predation level for Protozoa, Nematodes, and other microorganisms higher up in the predation order. With this predation more nutrients of all kinds are released into the nutrient pool around the root rhizosphere for consumption by the plant as well as by new bursts of diverse microbial life called upon to fulfill other soil health and plant protection maintenance tasks. In this instance it becomes obvious that the diversity of the microbial communities within a natural system must be extremely diverse and particularly if the concept of teamwork within microbial communities is considered, that it sometimes requires a team of different microbial communities to complete a task. The maintenance of these communities can therefore only be secured through a diversity of plants with each plant species responsible for the feeding of a different group because of its differing root microbial associative characteristics.
Nutrient feed sources from external organic material.
Organic debris such as leaves, and other decaying plant material also contribute to the nutrient pool as the complex carbohydrate molecules of cellulose, lignin and proteins are microbially degraded into sugars, amino acids, and elemental plant nutrients.
How does nature provide the other soil minerals, P, K, Ca, Mg, Fe, and others mentioned above?
In this regard there is a fungal group known as Mycorrhizae that perform a very important function in nature. There are about 260 known Mycorrhizae species in existence across 7 families and most certainly a large number not yet identified. They are also known to perform and fulfill their function working in teams of two or more species as well as with bacteria known as Mycorrhizae Helper Bacteria (MHB). These Mycorrhizae species are completely dependent on plants for their food though the root exudate source and the one group known as Vesicular Arbuscular Mycorrhizae (VAM) form root associated appendices known as arbuscules in host plant roots. Another group known as Ecto-Mycorrhizae colonize around the outside of the root tip within the rhizosphere.
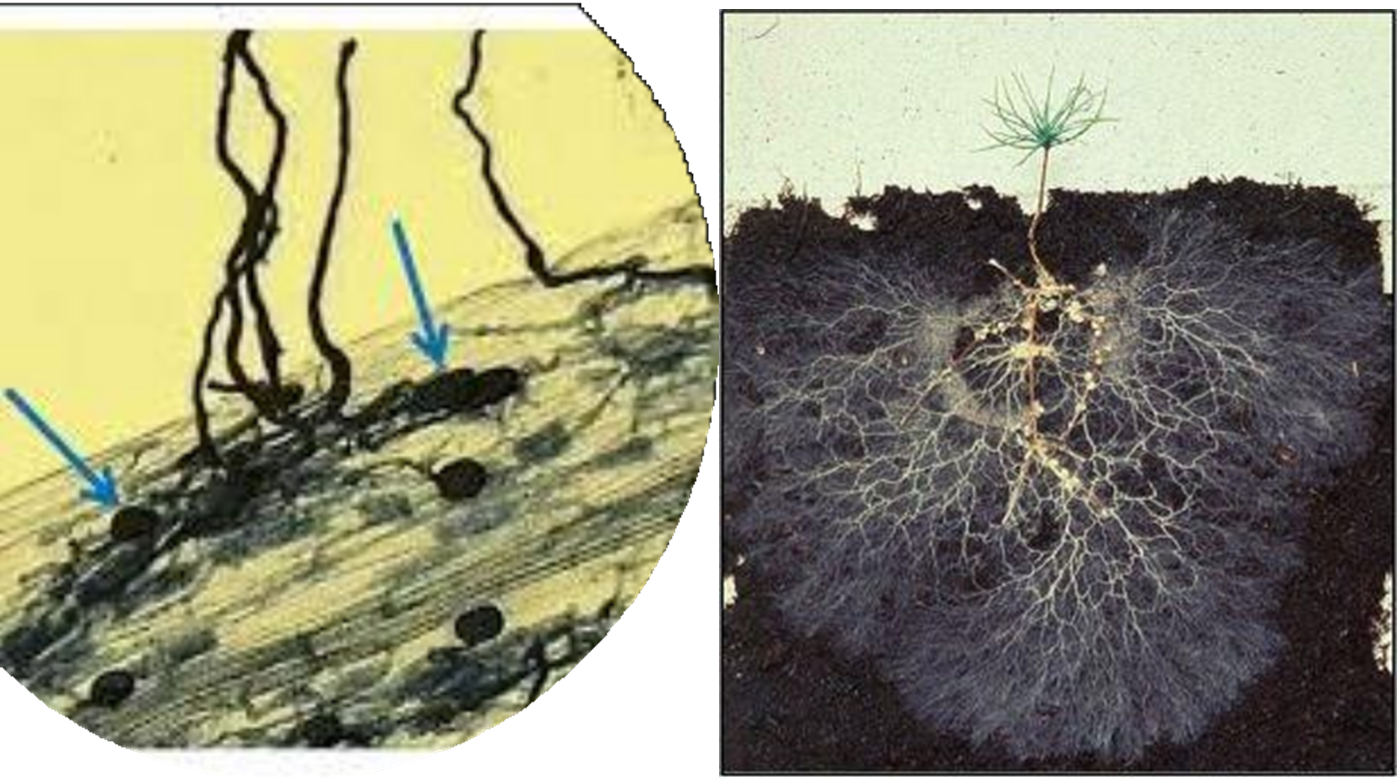
The pictures above indicate the arbuscules on the left picture and how Mycorrhizae hyphae can extend the root surface area on the right picture.
The fungal hyphae of the Mycorrhizae serve as root extensions adding miles of additional root transportation networks to the host plant. It is also a fraction of the thickness of a root hair and can penetrate hair line cracks to obtain nutrients as well as water for the host plant. Roots are unable to penetrate these micro pores where soil moisture is still available but not accessible to the roots and even when wilting point has technically been reached the Mycorrhizae hyphae can still extract water for the host plant.
Connectivity between plant and the soil mineral components.
The connectivity between host plant through Mycorrhizae attached or associated with the roots to the soil minerals is probably nature’s most important tool employed to provide the soil mineral components to the host plant. This connectivity also known as the “plant, root soil internet highway” that conveys inter-plant messages between plant and roots and also serves as the conduit for mineral supplies through the hyphae to the roots in an orderly fashion. The hyphal strands also connect plants within a community of the same and different species with each other creating an inter-plant communication system. This is a very interesting topic, presently receiving plenty of research attention confirming among others that an insect or other attack on one plant is signaled throughout the connected community so that natural defense mechanisms are triggered by the other community members in anticipation of attacks.
The fungal hyphae grow throughout the soil profile extending many meters beyond the host plants root systems attaching to many mineral sites where the different plant required elements are in abundance, however in the solid insoluble state. Should the host plant require any mineral to which the Mycorrhizae has an attachment to; a signal for such a request would be transmitted through the fungal communication highway to this site where the Helper Bacteria would solubilize the nutrient from the solid state and transport the required quantity back through the fungal hyphae to the root for use by the host plant where required in specific growth cycles. Fungal hyphae also excrete oxalic acid, one of the strongest organic acids that allow the hyphae to also “drill” their way through solid granite capturing elemental nutrients along the way.
In the process of hyphae extension an extremely concentrated organic carbon substance, with a Carbon content of more than 30%, known as glomalin is sequestered as a component of the hyphal membrane. Glomalin is a very stable, re-calcitrant glycoprotein, glue type molecule that can bind soil particles improving soil aggregate stability as segments of the Mycorrhizae hyphae senesce and become dysfunctional. The Mycorrhizal mutualistic association therefore not only provides nutrients to the host plants but also maintains other soil functions such as infiltration and water holding capacity through aggregation of soil particles.
The importance of this Mycorrhizae-aided nutrient supply system and its connectivity to the soil minerals cannot be over emphasized as it presents a system that naturally selects for the required nutrient as well as for the sufficiency thereof.
Plant root Mycorrhizal connectivity effect on plant, animal, and human health.
In Agricultural production systems where this Mycorrhizal connectivity is absent, as is the case with most conventional chemical-driven practices, plant roots are dependent on elements in the plant-available form in soil solution that its very limited root surface area contacts. The signaling system is largely impaired and roots can only absorb minerals when these become available within the vicinity of a root and are usually minerals from applied fertilizers or in the soil solution dissolved from the natural soil minerals at pH levels that solubilize these minerals through chemical means. The driver of conventional systems, where microbiology is largely absent or not taken into consideration, is therefore pH. The very important but non familiar micro and nano nutrients required for healthy individuals are therefore not readily available and thus not absorbed and become deficient within the food supply chain. The alarming issue here is that most new corn, wheat, soya beans and most other commercially grown crops are selected in breeding programs to accommodate these deficiencies and mainly selected for yield under higher and higher fertilizer regimes with narrower and narrower micro and nano nutrient requirements. The crops resulting from such breeding programs are therefore less nutrient dense with more carbohydrate bulk that supports the yield concept.
Rhizophagy is another natural microbial aided nutrient supply system very recently described in detail by Dr James White from Rutgers university; as a mechanism for the transfer of nutrients from symbiotic microbes (bacteria and fungi) to host plant roots that we term the ‘rhizophagy cycle.’ In the rhizophagy cycle, microbes alternate between a root intracellular endophytic phase and a free-living soil phase outside the root. Microbes acquire soil nutrients in the free-living soil phase within the rhizosphere as already described and then enter root tip meristem cells as complete bacteria cells to become endophytic, locating within the periplasmic spaces between cell wall and plasma membrane. Nutrients from these bacteria cells are then extracted through exposure to host-produced reactive oxygen in the intracellular endophytic phase that degrade the bacterial cell walls. Dr. White found that initially the symbiotic microbes grow on the rhizoplane in the exudate zone adjacent the root meristem outside the root. Inside the root tip the bacterial cells located within the periplasmic spaces of root cells and then convert to cell wall-less protoplast forms because of the reactive oxygen degrading of the cell walls effectively extracting nutrients from microbes. Surviving bacteria in root epidermal cells trigger root hair elongation and as hairs elongate bacteria exit at the hair tips, reforming their cell walls and cell shapes as these microbes emerge into the rhizosphere where they may obtain additional nutrients and the cycle is repeated. The rhizophagy cycle has been found to positively impact N, P, K as well as most macro- and micro-nutrients.
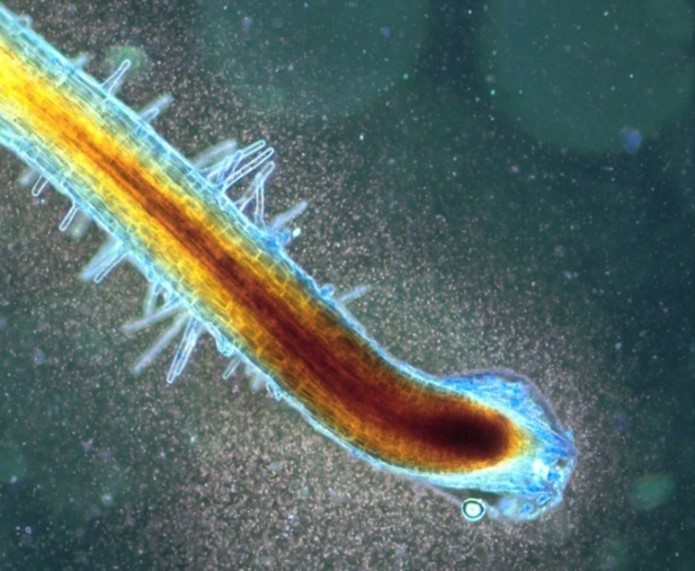
Image 1 is the root of Bermuda grass (Cynodon dactylon) with clouds of bacteria around the root tip exudate zone where microbes are entering into root cells.

Image 2 is the root tip root epidermal cells of curly dock (Rumex crispus) showing tetrads of the bacterium Micrococcus luteus within cells.
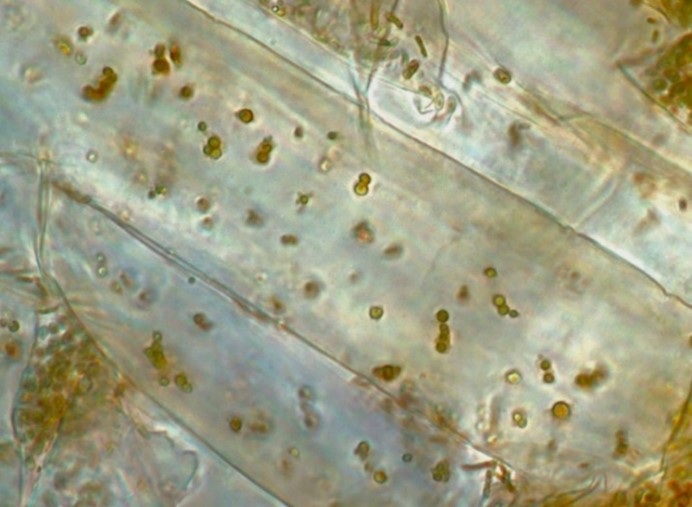
Image 3 is root epidermal cells of common reed grass (Phragmites australis) showing bacterial protoplasts replicating and degrading within the cell.
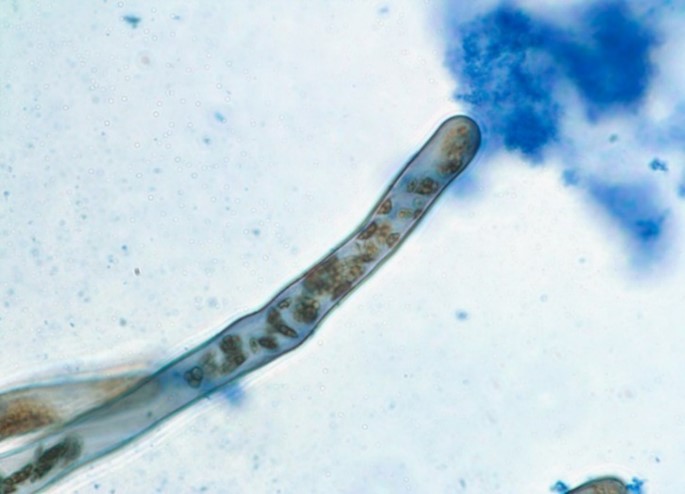
Image 4 is a root hair of a Fescue grass showing bacteria within the hair (brown due to hydrogen peroxide around bacteria) and bacteria ejected from the tip in mass (blue staining).
Concluding remarks.
From the above brief description of how natural systems, like forests, grasslands and other wild ecosystems survive, flourish and multiply, it is obvious that the natural resources providing the building blocks for the physical structure of all living entities is in abundance. The process of putting these building blocks together in a very orderly fashion to recreate and maintain healthy plants and animals that feed on these and ultimately humans that feed on both plants and animals is the complicated issue that has become a massive field of study collectively known as biology from where the Agricultural sciences have evolved.
Plants in a symbiotic association with soil microbes in the rhizophagy cycle; mycorrhizal associations as well as other types of microbial endophytes have many benefits to crop plants. These microbial associations increase the crop resistance of plants to oxidative stresses which translates to crops that are hardier and resistant to extremes in climate (heat, cold and drought) and other soil stresses such as salinity, heavy metals, and pathogens.
Crop science, as a discipline within the Agricultural sciences, is the scientific knowledge, learning, practice and understanding of the cultivation, management, processing and production of crops and how scientific principles can be applied to each of these. Unfortunately, this scientific discipline, be it the study of row crops referred to as Agronomy or perennial tree and vine crops referred to as Horticulture, have embraced chemistry rather than molecular biology in their endeavor to improve the production practices of these crops. The result is an ever-increasing divergence from the natural resource provision substituting this with chemistry aided production techniques substantiated through reductive research studies.
The consequences of this disregard for nature are more and more evident as we witness soil erosion, soil compaction, loss of soil organic carbon and producing crops with meaningless nutrition, albeit having huge yields.
Review Acknowledgement with thanks.
David C Johnson, Ph.D. Senior Research Scientist / Molecular Biologist Institute for Sustainable Agriculture, NMSU College Engineer
James F. White, Ph.D., Professor Department of Plant Biology Rutgers University New Brunswick, New Jersey


